
本能行为是与生俱来而无需后天学习的,对动物的生存与繁殖来说至关重要。单一本能行为,比如性、打斗、觅食和睡眠等,在果蝇和小鼠模型中均已取得了很多进展;但是,不同本能行为之间如何抉择,比如选择求偶而放弃睡眠或觅食,在神经机制上还知之甚少。有机体如何整合外界环境、内在生理状态以及过去的经验,在一系列复杂的价值比较后选择性输出某个特定的本能行为,是神经科学的一个根本问题。
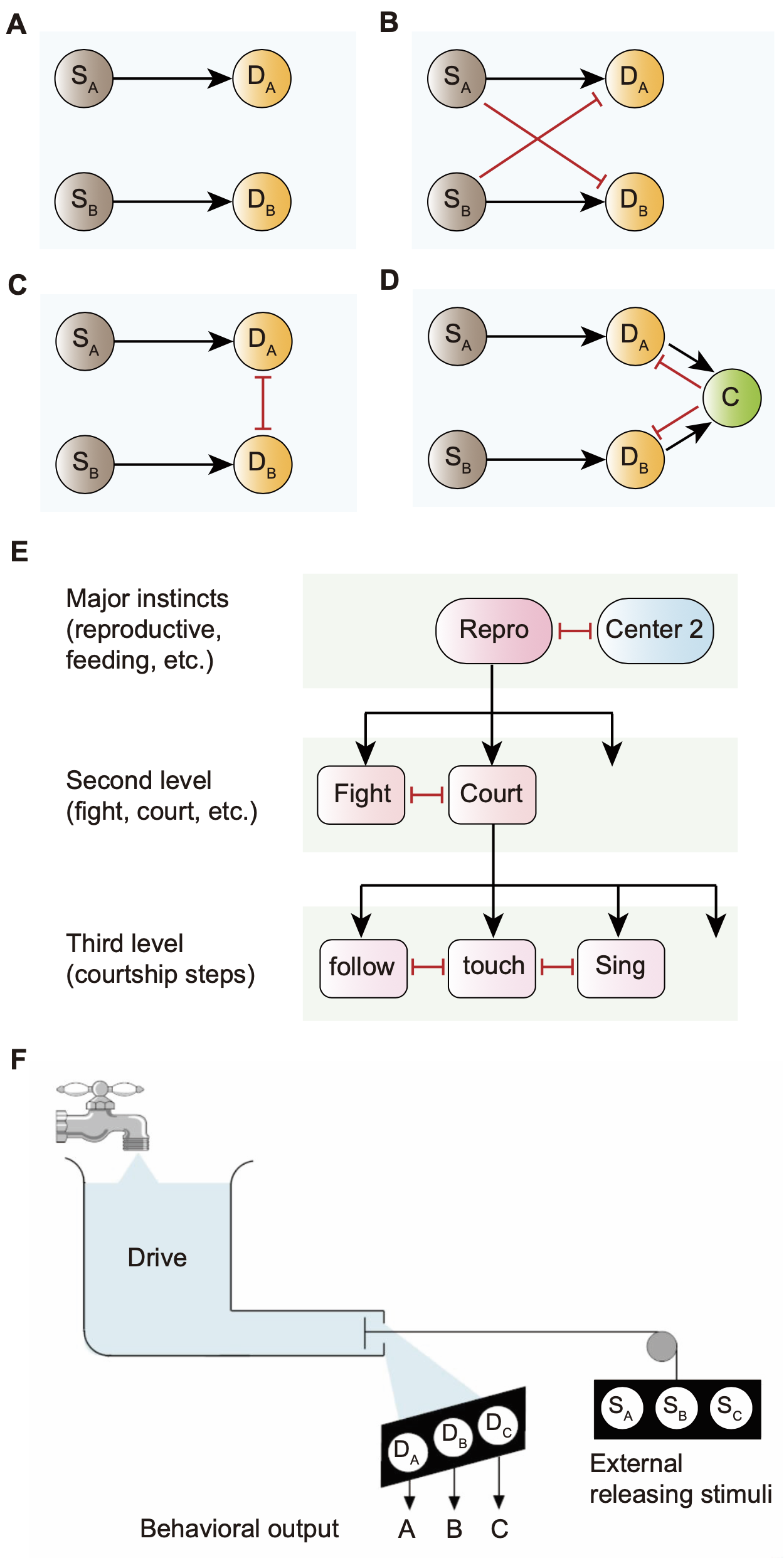
图1 行为抉择的计算模型示意图。(A-D) 从感觉神经元(S)到决策神经元(D)的线索积累通路(evidence-accumulating pathways)模型。SA和SB是接收外界信息的感觉神经元,DA和DB是整合线索的决策神经元。(A) 独立竞争模型。(B) 前馈抑制模型。(C) 决策神经元之间相互抑制的分布式模型。(D) 具有与其他决策神经元交互的公共神经节点的中央选择模型。黑色箭头表示兴奋,红线表示抑制。(E) Tinbergen 的等级模型。大脑中存在更高等级的决策中心,内外刺激共同作用于此。繁殖(repro)和觅食等主要的本能行为由不同的主要决策中心控制,高层级中心自上而下地调控低层级中心,每个层级的中心之间可能相互抑制。(F) Lorenz 的水力模型。内在状态就像流入水库的水提供压力(动力),而外界刺激的存在作为水阀释放压力并诱发某个行为。
本文首先总结了几类代表性的行为抉择模型(图1),包括诺贝尔奖获得者、神经行为学家Tinbergen和Lorenz分别提出的等级模型和水力模型;但是,这些模型都只能解释某些行为抉择过程中的特定现象,同时缺乏神经环路水平的直接证据。接着,本文以果蝇的性、打斗、觅食和睡眠这四种本能行为为例,总结了性相关行为与其他本能行为之间相互作用的神经环路机制,提出感觉信息的整合中心pC1神经元在本能行为抉择中可作为Tinbergen等级模型的“生殖中心”,并通过特定的方式与觅食和睡眠行为模块中的关键神经节点相互作用,以调控本能行为抉择(图2)。值得注意的是,pC1神经元具有较大的两性差异,其对性、打斗、觅食和睡眠行为的调控可能是本能行为及其选择性输出的两性差异的关键。在此基础上,本文提出了一个基于Tinbergen等级模型和Lorenz水力模型的混合模型,并在神经环路水平上解释了感觉信息如何整合并调控内在状态的“强度”和“持续性”,以及外界刺激如何在不同内在状态下诱发特定本能行为,从而产生行为抉择。
本文最后讨论了果蝇和小鼠模型中调控本能行为抉择的神经环路的相似性(图2),以及神经调质在行为选择或者转换过程中的重要作用。论文提出调控本能行为及其抉择的分子和神经环路在进化上具备一定保守性,各层次动物模型的研究将为理解高等动物包括人类的本能行为抉择提供一些共通的基本原理。
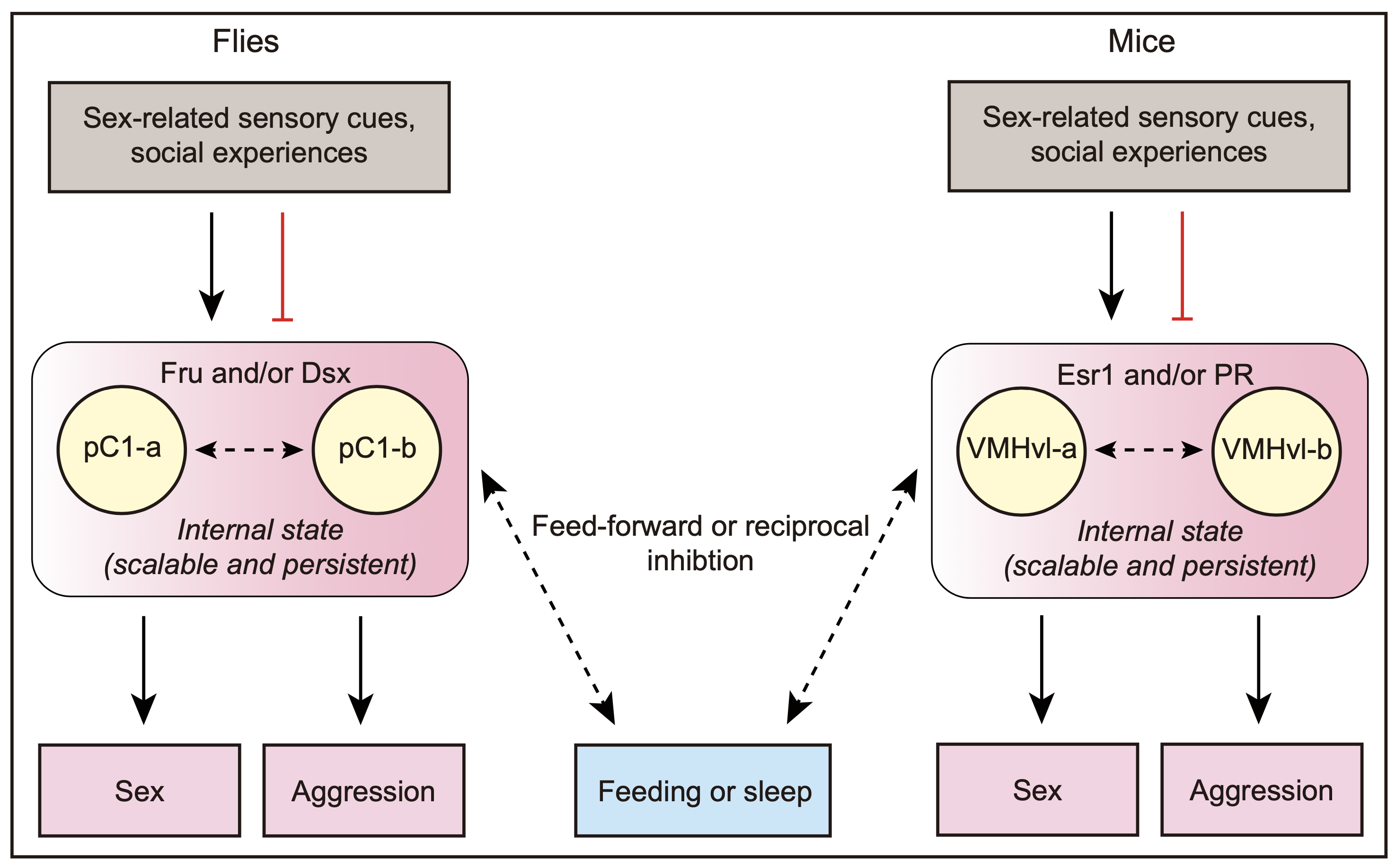
图2 雄性果蝇和小鼠中行为抉择的神经环路机制对比。果蝇中的pC1神经元和小鼠中的 VMHvl神经元都可以通过整合与性相关的感觉刺激和社交经验来编码生殖驱动。它们表达特定的性别调控因子,并且包括功能不同但关系密切的亚群,可直接调控性行为和打斗行为。它们还可以与其他本能行为的决策神经元交流,采用前馈或相互抑制等方式在生殖行为和其他竞争行为之间做出选择。pC1-a、pC1-b、VMHvl-a、VMHvl-b分别指代pC1和VMHvl神经元群体中不同的子集。黑色箭头表示兴奋,红线表示抑制,虚线表示尚不确定的连接。