
经典的孟德尔遗传定律告诉我们,一对等位基因将在减数分裂的过程中随着同源染色体的分离而分离,产生数量相同的两类配子,最后导致后代遗传到某一特定基因的概率为50%。但是自然界中的一些基因能够逃脱这个规则,以牺牲基因组中的其他基因为代价来增强自身的传播,这些基因就叫做自私的遗传元素(selfish genetic elements)。本来进化将偏好传播对个体有利的基因,但是自私的基因即使对个体无用甚至有害,都将无视进化的压力将其自身扩大到整个种群当中。
基因驱动(Gene drive)就是实现这种偏向遗传的系统,后代对于亲本的某个DNA序列的遗传将大于孟德尔定律的50%,通过有性繁殖,该序列在世代间传播的可能性被增强,最终在整个种群中成为优势序列。大多数工程基因驱动系统都是基于天然存在的自私的遗传元素的。所以也将自私的基因称为天然的基因驱动系统(natural gene drives),与实验室人工合成的基因驱动系统(synthetic gene drives)相对。当一个基因驱动修饰的个体和野生型个体交配后,理论上后代将全部遗传到亲本的基因驱动元件及其连锁的目的性状,这些新的后代又将“感染”更多的野生型个体,从而一代一代地产生级联放大效果(图1)。对于种群来说就将会出现两种结局:一种是种群崩溃或个体大幅度减少,这被称为种群抑制,相应的基因驱动类型就是“抑制型”;另一种是种群内的所有个体全都携带上目的性状,这被称为种群替代,相应的基因驱动类型就是“修饰型”。所以基因驱动是一种能够自我维持的、在种群水平上进行基因编辑的技术。基因驱动修饰的个体一经释放,就会席卷整个种群,直到地球上该物种中所有的个体都被改变,再无任何基因的交流(图1)。
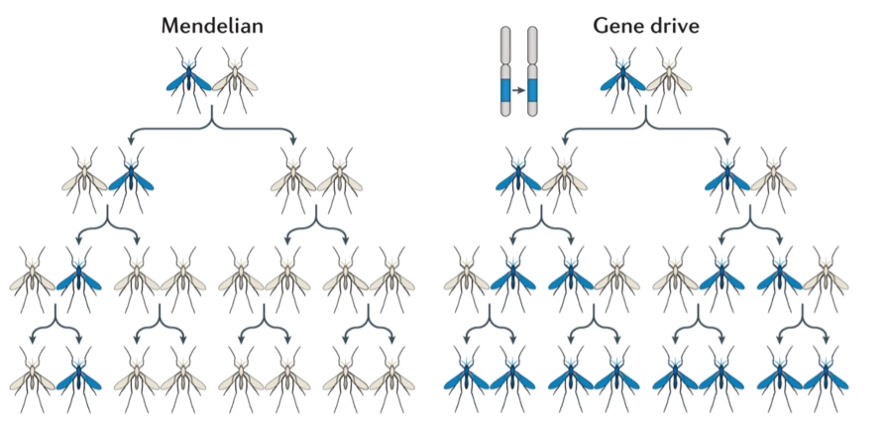
图 1 孟德尔遗传模式 VS 基因驱动遗传模式
由于基因驱动带来的种群抑制或是种群替代的两种结局,它能够被很好地应用来解决与公共健康和环境相关的生物学问题。如果传递的是降低物种适应性的性状,例如一些致死或者不孕不育的基因,那它可以用来根除疟疾、登革热等虫媒疾病,消灭或控制有害生物和入侵物种;但如果传递的是增加物种适应性的性状,则可以用来挽救濒危生物。在农业上还可以用祖先的等位基因替换后来产生的抗性等位基因来恢复其对杀虫剂或除草剂的敏感性。基因驱动方法的好处在于克服了进化上的劣势来传播有害性状,而对于有益性状的传播能比自然选择的更快也更彻底。
基因驱动技术发展了近一个世纪
基因驱动技术的基石——“自私的遗传元素”的现象早在一个世纪前就被观察到了。1928年俄罗斯遗传学家Gershenson发现野外捕捉到的雌性果蝇Drosophila obscura由于携带具有驱动效果的X染色体导致后代出现了雌性偏好的性别比例。1945年瑞典植物学家Gunnar Östergren在植物中发现了类似的现象,并首次明确地阐明了这种基因的自私性——“B染色体不需要对植物有益,它只需要对它自己有益就足够了”。大约在同一时间,其他几个自私的遗传元素的例子也被相继报道出来。但直到几十年后这个现象才得到广泛地关注。1980年,两篇关于自私基因的论文同期发表在Nature杂志上,向更广泛的科学界介绍了自私的基因的概念。这在科学界引起了非常激烈的讨论,随后的几期Nature杂志都成为了科学家们交流的场地。1988 年,John H. Werren和他的同事们就这个话题完成了第一篇综述,它创造了“自私的遗传元素”一词,并将其做了正式的定义,结束了令人困惑的术语多样化时代。2006年,帝国理工学院的科学家Austin Burt出版了第一本长篇书籍。一直已来对于自私基因在进化上的功能科学家们均持有不同的看法,但目前认为它在遗传中毫无作用除非从基因组冲突的角度来看待。
无独有偶,将现在被称为“基因驱动”的转基因生物释放到野外来解决公共健康和环境问题的思路早在1940年就被Serebrovsky提出了,随后Curtis在1968年进一步推广和阐明了该方法。两位科学家认为利用染色体相互易位(translocation)的杂合子个体半不育而纯合子个体可育的特性(图2),将人类所期望的性状连锁到易位的染色体上,随着繁殖易位染色体的频率将在种群中增加,如此就可以在目标害虫中传播目的性状从而达到控制害虫的目的。80年来,该议题都在持续地引发科学家们极大的研究兴趣。1994年,就着自私的遗传元素进入人们视野的东风,Hastings提出了可以利用该元件作为控制害虫的手段。虽然这种“基因驱动”的概念由来已久,但由于分子生物学上的技术限制,还没有人成功地在实验室里创造出来过。
转折点出现在10年后,2003年,“自私的遗传元素”领域内的专家 Austin Burt提供了第一个在实验室构建基因驱动的技术描述。他提出自然界存在的位点特异性的自私基因能将自己复制到目标DNA序列中,如果这些基因可以被设计为靶向新的目标序列,那么它们就可以用来操纵自然种群。至此,基因驱动技术正式由理论走向了现实。
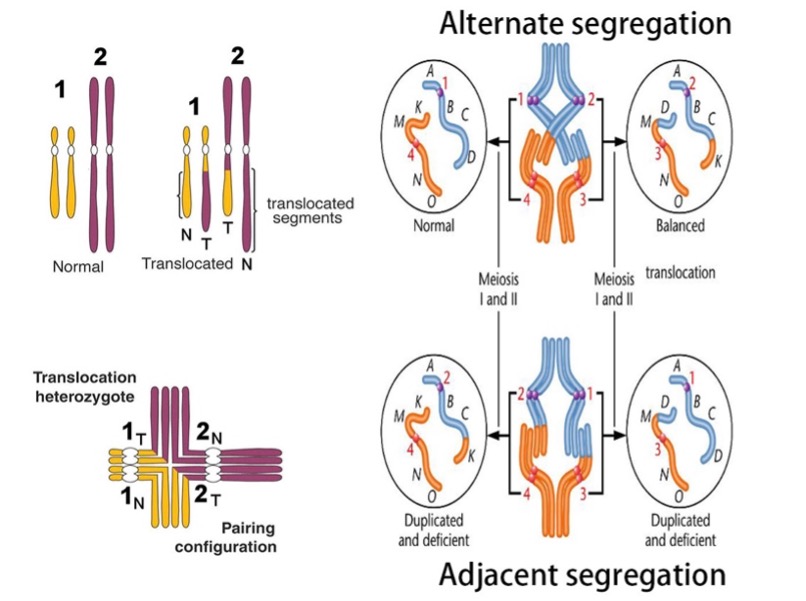
图 2 染色体相互倒位的杂合子半不育性
多样化的基因驱动系统
自私的遗传元素在促进自身传播的方式上是非常多样的,基于这些自然条件下存在的元件,研究员可以开发出具有不同特性的多种基因驱动类型。在评估针对特定应用的最适合的基因驱动类型和环境相关风险时,需要考虑以下几点重要的参数。
⑴、基因驱动传播的速度决定了野外初始释放的阈值。它除了与驱动的效率有关外,也与目标物种的交配动力学和产生后代的速度有关。传播速度高就认为该基因驱动类型具有很强的入侵性,所需要的初始释放阈值就比较低,否则就需要较高的释放阈值且更容易被局限在某个特定的区域内。
⑵、为了遏制非目的的传播,基因驱动对于所针对的物种必须是特异的。
⑶、基因驱动在较长的一段时间内存在并保持活性的目的要求它们在进化上是稳定的,所以基因驱动应该具有较低的适应性成本。
⑷、在基因驱动传播的过程中,目标物种很可能会产生针对驱动的抗性。这些抗性基因可以是本身就存在的,也可以是响应驱动而产生的。所以维持抗性的产生处于一个较低的速度是研究人员们所期望的,除非想利用抗性将基因驱动限制在较小的种群中。
⑸、最后,基因驱动对生态的破坏是未知且彻底的,所以基因驱动需要是可逆的,甚至是可移除的。对于一些传播速度低的驱动类型,可以通过向野外释放大量的野生型生物将其从种群中完全移除;其他的类型可以通过构建第二代基因驱动进行恢复,但要注意的是,逆转了基因驱动的生物,不代表能逆转其对生态环境的影响。
最有效的基因驱动系统——Homing drives
归巢核酸内切酶基因(HEG)为构建和测试合成基因驱动系统提供了第一个实用工具。Homing 也就是归巢是指将自身复制到相对的同源染色体等位基因上的过程。自然界中存在250多种归巢核酸内切酶基因,他们可以特异性地识别并切断与它们相对的同源染色体上的等位基因,从而迫使细胞快速修复这种DNA断裂。断裂后,细胞可以使用HEG作为同源重组修复的模板,导致HEG将自身复制到其竞争的等位基因中。但是该方法的难点在于让天然存在的HEG去识别那些不是它的先天识别序列的新序列,这个过程需要复杂的蛋白质工程,并且已经被证明非常困难。也就是说,不是任何的DNA序列都可以被操纵。
这个问题可以被可编码的核酸酶,例如锌指核酸酶(ZFNs)和转录激活物样效应核酸酶(TALENs)绕过,二者开启了靶向基因组编辑的时代。然而锌指和TALENs虽然能被设计来切割新的靶序列,但是他们大量的重复序列之间很可能会发生同源重组导致他们在进化上不稳定。在一次复制事件后,每个驱动分别只有75%和40%足够完整能够催化第二轮的复制,随着时间的推移,基因驱动的效率将会大打折扣。
CRISPR-Cas9细菌防御系统的发现和2012年Doudna团队开发的简化双组分系统,包括Cas9内切酶和一种易于编码的引导RNA (gRNA),几乎彻底改变了生物学的所有领域。易于编码的gRNA使有效地切割任意DNA序列成为可能;Cas9酶在多数物种中保持的高酶活性使基因编辑不再只局限于模式生物中;二者低重复的结构使其在进化上更为稳定,能够长久地发挥作用。据此,2014年发表了综述“Concerning RNA-guided gene drives for the alteration of wild populations”讨论了将CRISPR–Cas9技术作为构建合成基因驱动系统的普遍手段的可能性。CRISPR-Cas9系统的两部分性质和灵活的可编码性导致了细菌、昆虫、甚至哺乳动物中各种基因驱动系统的快速发展(图 3)。
在新的结构中,Cas9酶替代了自然存在的归巢核酸内切酶,gRNA被设计为靶向任意序列,二者连锁在一起同时复制到同源染色体中的目标位点上实现了基因驱动自我维持的特性。当基因驱动的染色体和野生型的染色体相遇后,将发生以下3种可能:①是没有发生切割,这可以通过增加gRNA的数量来增加切割的效率;②是切割发生后,通过非同源重组的末端连接(NHEJ)进行修复,目标序列中核苷酸的插入或缺失将产生一个抗性等位基因并进一步阻止它被核酸内切酶识别和切割。这些抗性等位基因在与基因驱动的对抗中幸存下来后将受到进化的青睐,随着大量增加最终消除基因驱动;③是切割发生后,通过同源重组进行修复(HDR),实现我们所期望的基因盒复制。所以为了基因驱动的效率,最小化NHEJ以及最大化HDR是研究员们所努力的方向。对抗抗性位点最好的手段是靶向多个位点,只要切割效率够高,在所有位点积累突变就会变得困难,而CRISPR-Cas9系统可以很简单的实现这一点。此外,采用Cas9切口酶配合一对gRNA,分别在2条DNA链上产生小切口,从而形成功能性的双链断裂,也能增强识别特异性以及同源重组修复的概率。因此Homing drives的传播速度是最快的,但同时它产生抗性的速度也是最快的。
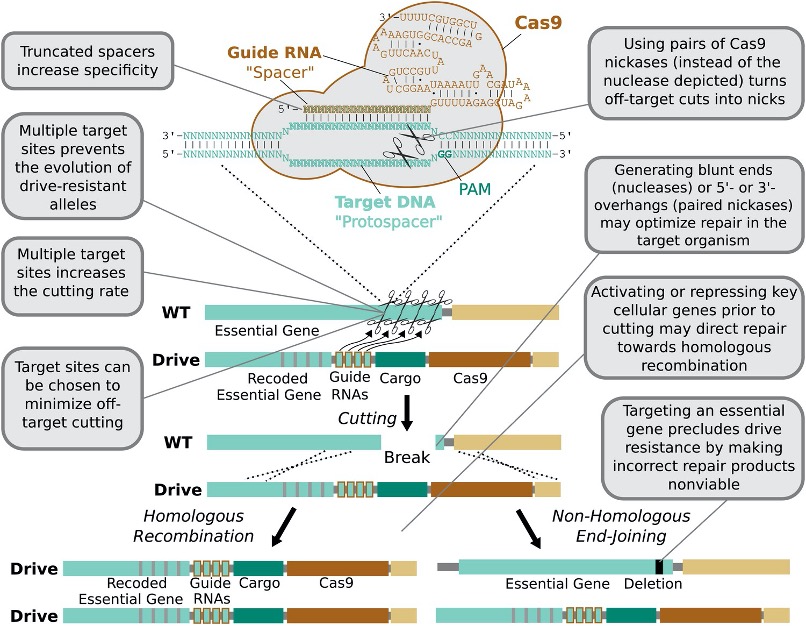
图 3 CRISPR-Cas9系统应用于基因驱动中的技术优势
Homing drives的有效不仅体现在它极高的效率上,还体现在它可以被应用于种群抑制和种群修饰这两大基因驱动类型中。如果gRNA针对的是对生存繁殖必不可少的重要基因,gRNA–Cas9盒的插入将会导致基因功能的丢失,造成纯合致死或不孕不育的结果。一旦大部分群体成为杂合子,就会导致种群抑制,因为杂合子配对会产生致死或无法繁殖的后代。而如果在gRNA–Cas9盒中再连锁上某个基因,就会导致目的基因随着gRNA–Cas9盒的复制而在种群中传播,最终实现种群的修饰。
“抑制型”基因驱动的主要挑战就是驱动可能无法实现其抑制的目的。随着世代的增加,基因驱动的频率在种群中上升的同时,该种群的平均适应性也会随之降低,更不要提还可能会产生的功能性抗切割的等位基因。这些抗抑制驱动的性状将被进化积极地选择,导致抑制驱力的消失甚至种群的反弹。产生适应性成本的代际延迟将增加驱动的抑制能力,例如,无孙子>不育>致死表型。“抑制型”基因驱动就像杀虫剂一样,可能需要在同一地点反复使用,随着领域的扩大,将造成不断升级的成本和后勤保障难度。另外,在当地消灭一种物种可能导致其他物种填补了空的生态位,产生意想不到的生态后果。“修饰型”基因驱动相对于“抑制型”的一个显著的理论优势在于预计它们可以在种群中保持足够长一段时间(2-5年)的稳定,以能够有充足的时间达成目的。研究员们可以将驱动设计为不携带适应性成本,如果有的话成本也可以是最小的。而且,由于“修饰型”没有留下任何空白的生态位,可能施加更少的环境影响。
实现种群抑制的基因驱动系统——Sex-linked meiotic drives
一些自私的遗传元素通过操纵遗传传递过程来达到自己的目的。在减数分裂过程中,本应平等分开的等位基因的传播出现偏离,最终在配子中“过度存在”。这种扭曲可以以各种方式发生,被统称为分离扭曲(segregation distortion)。比如有些元素可以优先在卵细胞而不是极体中传递,只有前者会受精并将遗传物质传递给下一代。不过许多形式的分离扭曲发生在雄性配子形成的过程中,在精子成熟或精子发生的过程中,携带不同等位基因的精子细胞具有不同的死亡率。任何能够控制自己最终进入卵子而不是极体或是在生殖细胞成熟过程中成功存活下来的基因都将具有传播优势,并能够在种群中增加它们的频率。当减数分裂驱动作用于性染色体时,它们就会扭曲后代的性别比例。
构建在Y染色体上的X染色体粉碎机就能够使后代出现雄性偏好的性别比例。Y染色体上携带了Cas9酶和针对多个重要位点的gRNA,在精子发生过程中Cas9酶将靶向并粉碎X染色体,从而确保只有携带Y染色体的精子才有活力。理论上当修饰过的雄性(XY*)与野生型的雌性(XX)交配后,后代将全部为雄性,而且这些继承修饰过的Y染色体的雄性能够继续发挥作用,使种群中雄性持续增加而雌性持续减少,最终导致种群崩溃(图 4)。Sex-linked meiotic drives将用于实现种群抑制,它们的传播速度适中,但产生抗性的速度很低。
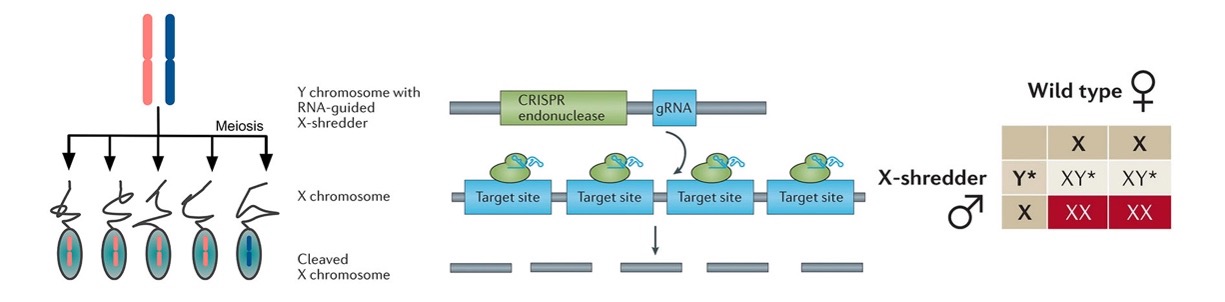
图 4 Sex-linked meiotic drives的原理、结构和遗传模式
实现种群替代的基因驱动系统——Medea drives & Underdominance drives
Medea全称maternal effect dominant embryonic arrest,该类自私的遗传元素通过诱导那些没有从亲本遗传到Medea因子的后代产生母本效应致死而让自身存活下来。由于Medea因子在适应性上的优势,如果将目标基因连锁到该因子上,那么荷载的基因就能够在种群中存活并传播。
自然界中存在的Medea的分子基础还不清楚,人为设计的Medea基因驱动系统可以通过毒素-解毒剂的配合来实现。早先采用的毒素是microRNA,通过RNA干扰来破坏重要的胚胎基因,它们在后续的个体发育过程中起到关键的作用,基因功能的丧失将导致个体的死亡,即使从父本那得到一条野生型的染色体也不能阻止这个结果的发生。在CRISPR-Cas9技术出现后,毒素变为效率更高的gRNA,通过切割后的非同源末端连接修复产生基因突变导致功能缺失。解毒剂是针对毒素设计的因子,实验室里可以被设计为相应的被挽回的破坏基因本身,但是这个外源的基因要重新编码以免疫gRNA的识别。Medea驱动的关键在于,毒素和解毒剂在完全错开的两个时期表达。毒素表达于雌性的生殖细胞中,破坏所有胚胎中的胚胎必需基因。而解毒剂表达于早期胚胎的合子阶段,只有那些遗传到Medea因子——胚胎必需基因被挽回了的胚胎能够存活下来。所以携带Medea的杂合子雌性与野生型雄性交配后,只有50%的后代能够存活下来;当然父本的Medea因子也能发挥挽回的作用,杂合的Medea雌性与同样杂合的Medea雄性交配后将有75%的后代存活(图 5)。
那些要求转基因的频率超过某个阈值才能成功传播的基因驱动是非常有吸引力的,因为它们带来的是局部而不是全局的替换,并且转基因可以通过释放大量野生型个体来消除。当携带转基因的染色体相比起非携带转基因的同源染色体经历频率依赖的适应性改变时,(例如前者在低频时具有较低的适应性而在高频时具有较高的适应性),阈值依赖的基因驱动就可以发生。这类在有效驱动和失控之间找到了一个平衡的基因驱动就叫underdominance drives。在种群遗传学中,显形不足(underdominance)是一种逆导致基因型多样性的种群分布倾向的选择,也可以被称为杂合致死(heterozygote inferiority),是指具有杂合子基因型的二倍体个体的适应性低于相应的纯合子基因型。如果杂合子的适应性小于两个纯合子的适应性,就可以认为两个等位基因彼此之间是显性不足的。该系统表现为一个双稳态开关模式,也就是说一个种群被期望处于两种等位基因频率的稳定状态中的任意一个。当转基因出现的频率高于阈值频率时,这个基因将传播到整个种群中,而当它们的频率低于该阈值时,则从种群中丢失。
大多数阈值依赖的基因驱动尚未实现。虽然染色体相互易位就是一个典型的例子,但是其巨大的适应性成本和将感兴趣的基因紧密连锁到易位断点上的困难,阻碍了这类方法的进一步发展。目前在实验室中开发了配合的双位点毒素-解毒剂的underdominance系统,也可以被称为母本效应致死的underdominance系统。它的原理和Medea一致,差别在于采用了两种不同的母本表达毒素,而每种毒素确实都与一种合子期表达的解毒剂相连,只不过解毒剂针对的是另一种毒素的,所以只有两条染色体都存在时,后代才能存活(图 5)。两类毒素-解毒剂的结构(A & B)可以位于一对同源染色体的同一位置(单位点UDMEL)。在这种配置中,可存活的后代总是异型结合的杂合子(transheterozygotes),携带两个不同的突变等位基因(A/B),每一代有一半的后代死亡。另外,该结构也可以位于2条不同的非同源染色体上(双位点UDMEL),在这种情况下,将有更多的基因型存活下来(AABB;A+B+)。根据模型预测,在没有其他适应性成本的条件下,单位点系统要求transheterozygotes雄性的释放阈值为总种群的78%;而双位点常染色体UDMEL对于释放双纯合子的雄性将要求更低的阈值,为24%。具有额外适应性成本的元素仍然可以驱动,只不过它们的释放阈值将会有所增加。此外,值得注意的是,单位点UDMEL具有这样一种特性:当替换的转基因固定到种群中时,重复引入野生型雄性将会导致被替换的群体的抑制。这是因为UDMEL雌性(A/B)和野生型雄性(+)之间的交配由于孟德尔遗传定律只能产生带有错误解毒剂的后代,因此它们是不可存活的。当然如果抑制之后重新引入野生型的雄雌个体,种群数量是会恢复的,而转基因由于它们的频率低于驱动所需的频率而丢失。
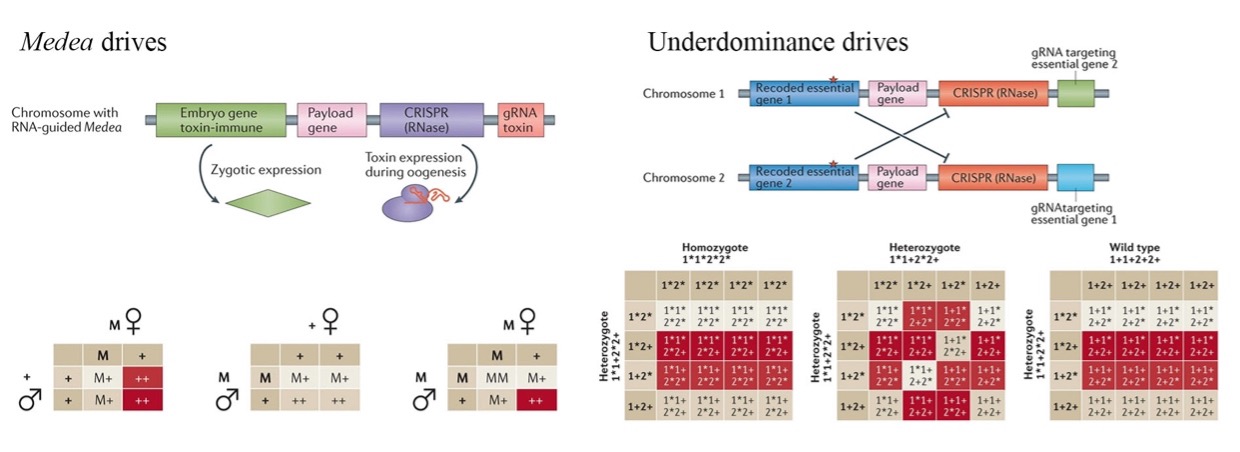
图 5 Medea drives和Underdominance drives的结构和遗传模式
Medea drives和Underdominance drives都将用于实现种群替代。相比于通过直接修改等位基因来增加荷载基因的频率,它们二者都是通过死亡一部分后代造成的适应性优势来达到传播的目的的,所以它们的传播速度都将低于homing drives。Medea drives的传播速度适中,但产生抗性的速度很低。而Underdominance drives的传播速度是最低的同时还有较高的产生抗性的速度,所以该类型驱动能够被很好的局限在某一区域内,并且可以通过向野外释放大量的野生型个体来将其彻底地移除。这些基因驱动系统或多或少具有不同的限制性和可逆转性,比如单位点UDMEL的可控性>常染色体-常染色体双位点UDMEL>Medea,可以用于创建一系列梯度基因驱动系统适用于特定的社会、监管和物理环境。
采用基因驱动技术需要慎之又慎
基因驱动修饰的生物确实能产生巨大的利益,但是由于它的快速传播,自我维持以及不可逆转的特性导致危害生态的潜力也是巨大的。比如,在当地消灭一种物种可能导致其他物种填补了空的生态位,产生意想不到的生态后果;基因驱动的脱靶效应可能将其传播到预期外的地区甚至通过罕见的交配事件穿越到另一个物种中;基因驱动还可能被滥用造成经济损失甚至是生物恐怖主义。更重要的是在伦理上,一个物种是否应该被修改甚至是被消灭是能够由人类所决定的吗?因此,基因驱动不再仅仅是一个科学问题,它还将涉及到伦理、法律和社会方面。
2016年,美国国立卫生研究院 (NIH) 及其基金会要求美国国家科学、工程和医学研究院(NASEM)召集一个专家小组,就“我们对基因驱动了解多少”这一主题进行独立、客观的评估,并总结成了一本书——《基因驱动正当时》。虽然没有足够的证据支持将基因驱动改良的生物释放到环境中,但该小组建议应继续进行基因驱动的研究,因为基因驱动对基础和应用的好处是显而易见的。委员会提出每个基因驱动的研究在开始前都应进行充分的思考和独立的科学评估,包括它实现预期目标的能力,对其他物种的可能影响,对生态系统和人类社会的影响以及释放后的挽回措施。从实验室研究到最终释放到野外的过程中还应增加受控的区域测试,安全应该是每个阶段的主要考虑因素。同时,针对基因驱动开发的监管也要同步落实,尽管没有专门针对基因驱动的立法,但它们的使用需要当地的同意,并且公众需要及早且透明的了解到整个过程。
对于研究者来说,遵守在实验室进行基因驱动的操作规范更是必要的。2015年7月出版的Science杂志发表了数位资深的基因驱动研究员撰写的关于在实验室进行基因驱动研究的安全指南。他们指出未来的研究应尽可能地结合使用这些严格的限制措施,并始终使用以防止基因驱动修饰生物无意识地释放到自然种群中。这些限制措施可以总结为分子、生态以及物理屏障这三个方面。在分子层面上,设计基因驱动方案的时候,研究员们可以考虑将Cas9酶和gRNA分开,基因驱动仅当与未连接的Cas9酶结合时才能传播,这样不小心泄露的转基因生物与野生型个体交配时,驱动力的基本要求没有得到满足;或者预先构建一个驱动系统传播对种群没有影响的序列,再将具有功能的驱动设计为针对这些野外物种所缺乏的合成序列也是防止泄露造成恶性结果的好办法。在生态层面上,研究员可以在不是目的物种的栖息地或缺乏野外潜在交配对象的区域进行实验。最后,基因驱动修饰生物和外界环境之间的物理屏障是必不可少的,只有在转基因生物完全失活后才可以移开屏障。
by 蒋昕钰